Legume symbioses
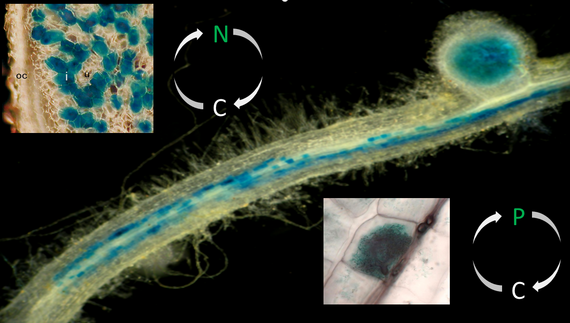
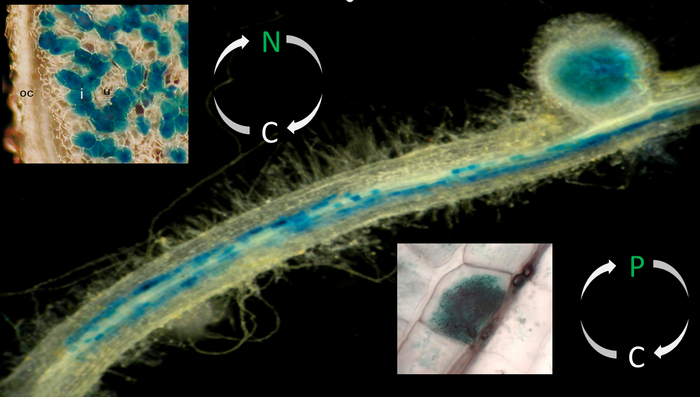
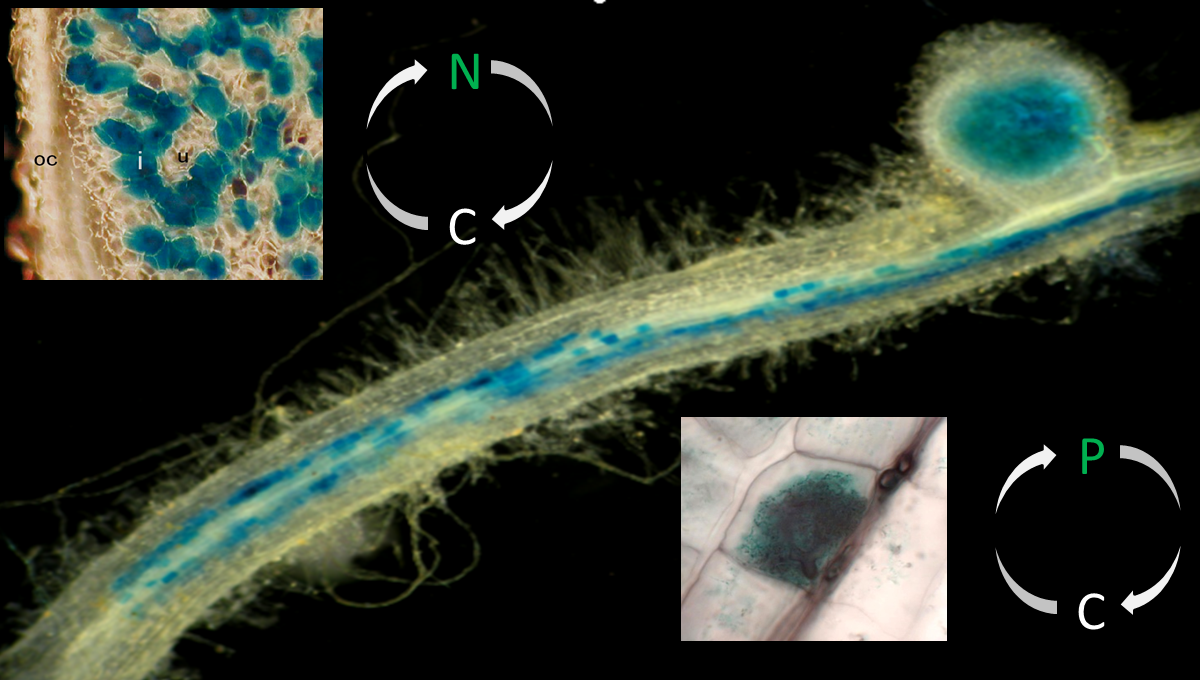
Legume plants enter symbiotic interactions with beneficial soil microbes. Whereas the interaction with rhizobial prokaryotes leads to the formation of nitrogen-fixing root nodules, the colonization of roots by soil fungi of the phylum Glomeromycota results in the establishment of an arbuscular mycorrhiza that improves the plant's phosphate uptake. In return for supplying limiting nutrients, the microbial symbionts obtain carbon compounds from the plant.
Root nodules
A remarkable feature of legumes is their capacity to enter a nitrogen-fixing root nodule endosymbiosis with rhizobial prokaryotes. This extremely host-specific interaction is initiated in the root cortex after a perception of species-specific rhizobial lipochitooligosaccharide (LCO) nodulation factors (Nod-factors) through membrane-bound LysM receptor kinases. Signal transduction triggers the expression of additional genes located further downstream in the Nod-factor perception pathway and finally results in the organogenesis of a unique plant organ, the root nodule.
Nodules are colonized by the rhizobial prokaryotes through intracellular infection threads. Subsequent to the uptake of rhizobia at the infection thread tips, cells from the inner region of root nodules house the nitrogen-fixing endosymbiotic microsymbionts in transient organelles referred to as symbiosomes. Here, rhizobia differentiate into bacteroids capable of carrying out biological nitrogen fixation, the conversion of atmospheric dinitrogen to ammonia. To drive bacteroid energy metabolism required for the cleavage of dinitrogen, the plant supplies dicarboxylic acids to the endosymbiotic prokaryotes. In exchange, ammonia is released through channel proteins located in a highly specialized perisymbiotic membrane.
Indeterminate root nodules e.g. formed by legumes of the genera Vicia, Pisum, and Medicago are characterized by a persistent apical meristem that leads to the continuous formation of zones in the central nodule tissue: the meristematic zone, the prefixing zone, the interzone, the fixation zone, and the senescence zone. Apart from studying nitrogen fixation, the formation of indeterminate root nodules is an excellent model to address processes of general relevance for plant biology, e.g. the de-differentiation of root cortical cells prior to nodule primordium establishment, the formation of a persistent apical meristem, the differentiation of specific tissues, and the sharing of metabolic tasks between infected and non-infected cells.
Arbuscular mycorrhiza
Whereas the root nodule symbiosis is unique to legumes, more than 80% of higher plants enter an arbuscular mycorrhiza (AM) endosymbiosis with fungi of the phylum Glomeromycota, with prominent representatives being Rhizophagus irregularis (formerly Glomus intraradices) and Glomus mosseae. Following a pre-symbiotic signal exchange that involves fungal (lipo)chitooligosaccharides (COs/LCOs) similar to rhizobial Nod-factors, hyphae from the extraradical AM fungal mycelium penetrate the root epidermis through a hyphopodium and subsequently proliferate in the inner cortex. These intraradical, intercellular hyphae terminate in highly branched, intracellular structures designated arbuscules.
Comparable to root nodules, nutrient exchange occurs at the intracellular interface between the symbionts. Here, a periarbuscular membrane and a periarbuscular space surrounds the fungal arbuscules. In return for the supply with carbon compounds (sugars and fatty acids), the fungal microsymbiont in particular transfers phosphorus, but also nitrogen, minerals, and water to the plant. Arbuscules are the major sites for nutrient uptake by the plant, with mycorrhiza-specific nutrient transporters being exclusively localized at the arbuscular interface.
From a functional point of view, AM essentially enlarges the size and surface area of a plant root system that way facilitating an optimized uptake of nutrients from the soil. Due to the emergence of arbuscular mycorrhiza more than 400 million years ago, it is speculated that the ability to enter this symbiosis was a prerequisite for the evolution of land plants.





